Energia muundamine kloroplastides. Kuidas päikesevalguse energia muundub III peatükk
On laialt teada tõsiasi, et Päike on taevakeha (täht) ja päikeseenergia on sisuliselt tema elulise tegevuse tulemus. Sellel toimuvad protsessid eraldavad tohutul hulgal energiat, paiskades selle uskumatu kiirusega meie planeedi poole. Päikeseenergia kasutamine juhtub inimestega nii teadlikult kui ka alateadlikult. Päikesekiirtes supeldes ei mõtle me sellele, et selle tähe energia käivitab meie kehas mitmeid olulisi protsesse (näiteks D-vitamiini toodetakse meie nahas); tänu sellele toimub taimedes fotosüntees; Veeringe looduses on ka "tema töö". Me võtame seda iseenesestmõistetavana. Kuid see on vaid osa päikeseenergia rollist meie elus.
Päikeseenergia praktiline kasutamine
Kõige lihtsam ja kõigile tuttavam päikeseenergia kasutusviisid- selle kasutamine kaasaegsetes kalkulaatorites (väga kompaktsetel päikesepaneelidel) ja majapidamisvajaduste jaoks (kuivatatud puuviljad, vee soojendamine maal asuva väliduši paagis). Päikese soojusest soojendatud õhu liikumine tagab ventilatsioonisüsteemi ja korstnate töö. Päikesekiiri kasutatakse merevee magestamise aurustajana. Päike on üks peamisi energiaallikaid satelliitide pikaajaliseks tööks, samuti kosmoseuuringuteks kasutatavate seadmete jaoks. Meie ellu tuuakse üha enam elektrienergial töötavaid autosid.
Päikeseenergia vastuvõtmine ja muundamine
Päikeseenergia tabab meie planeeti kolme tüüpi kiirguslainetena: ultraviolett-, valgus- ja infrapunalainetena.
Päikeseenergia kasutamine peamiselt soojuse või elektri tootmiseks. Infrapunalained, mis langevad teadlaste välja töötatud spetsiaalsele pinnale, muutuvad meile vajalikuks.
Seega kasutatakse soojuse eraldamiseks kollektorit, mis neelab infrapunalaineid, salvestusseadet, mis seda akumuleerib, ja soojusvahetit, milles toimub kuumutamine.
Elektrienergia tootmisel kasutatakse spetsiaalseid fotoelemente. Need neelavad valguskiiri ja vastavad paigaldised muudavad need kiired elektriks.
Päikeseenergia kasutamise viisid saab jagada sõltuvalt elektrijaama tüübist selle töötlemiseks. Kokku on neid kuus.
Esimesed kolm: torn (disain musta torni kujul, mille sees on vesi ja ümber peeglid), paraboolne (meenuvad sees peeglitega satelliiditaldrikud), taldrikukujuline (näeb välja nagu peeglitest lehtedega metallpuu). Neid saab kombineerida, kuna neil on sama tööpõhimõte: nad püüavad kinni teatud koguse valgust, suunavad selle vedelikumahutisse, mis kuumeneb ja vabastab auru, mida omakorda kasutatakse elektri tootmiseks.
Neljandaks- fotosilmadega varustus. Kõige kuulsam tüüp, kuna selle mõõtmed võivad sõltuvalt vajadusest erineda. Väikeseid päikesepaneele kasutatakse eramajapidamiste vajadusteks, suuremaid tööstuslikeks vajadusteks. Toimimispõhimõte seisneb selles, et fotoelemendis neeldunud päikesekiirtest toota elektrit selle sees oleva potentsiaalide erinevuse tõttu.
Viiendaks- vaakum. Struktuurselt on tegemist ümmarguse klaaskatusega kaetud maatükiga, mille sees on turbiinidega torn jaluses. Tööpõhimõte on selle katuse all maapinna soojendamine ja õhutõmbe tekitamine temperatuuride erinevuse tõttu. Turbiini labad pöörlevad ja toodavad energiat.
Paljud meist on ühel või teisel viisil päikesepatareidega kokku puutunud. Keegi on kasutanud või kasutab päikesepaneele koduseks elektri tootmiseks, keegi kasutab väikest päikesepaneeli oma lemmikvidina laadimiseks põllul ja kindlasti on keegi näinud väikest päikesepatarei mikrokalkulaatoril. Mõnel oli isegi õnn külla tulla.
Kuid kas olete kunagi mõelnud, kuidas toimub päikeseenergia muundamine elektrienergiaks? Milline füüsikaline nähtus on kõigi nende päikesepatareide töö aluseks? Pöördume füüsika poole ja mõistame genereerimisprotsessi üksikasjalikult.
Algusest peale on ilmne, et energiaallikaks on siin päikesevalgus ehk teaduslikult öeldes saadakse see tänu päikesekiirguse footonitele. Neid footoneid võib ette kujutada Päikeselt pidevalt liikuvate elementaarosakeste voona, millest igaühel on energiat ja seetõttu kannab kogu valgusvoog mingisugust energiat.
Päikese pinna igalt ruutmeetrilt eraldub kiirgusena pidevalt 63 MW energiat! Selle kiirguse maksimaalne intensiivsus jääb nähtava spektri vahemikku - .
Niisiis on teadlased kindlaks teinud, et päikesevalguse voolu energiatihedus Päikesest Maale 149 600 000 kilomeetri kaugusel pärast atmosfääri läbimist ja meie planeedi pinnale jõudmist on keskmiselt umbes 900 W ruutmeetri kohta.
Siin saab seda energiat vastu võtta ja püüda saada sellest elektrit ehk teisendada Päikese valgusvoo energia liikuvate laetud osakeste energiaks ehk teisisõnu selleks.

Valguse muundamiseks elektriks vajame fotoelektriline muundur. Sellised muundurid on väga levinud, need on saadaval tasuta müügiks, need on nn päikesepatareid - fotoelektrilised muundurid ränist välja lõigatud vahvlite kujul.
Parimad on monokristallilised, nende kasutegur on umbes 18%, see tähendab, et kui päikesest lähtuva footonivoo energiatihedus on 900 W/m², siis saab arvestada 160 W elektrienergiaga ruutmeetri kohta. sellistest elementidest kokku pandud aku.
Siin töötab nähtus, mida nimetatakse "fotoefektiks". Fotoelektriline efekt või fotoelektriline efekt- see on nähtus, mille käigus toimub aine elektronide emissioon (nähtus, et elektronid paiskuvad välja aine aatomitest) valguse või muu elektromagnetilise kiirguse mõjul.
1900. aastal tegi kvantfüüsika isa Max Planck ettepaneku, et valgus kiirgub ja neeldub üksikute osade või kvantidena, mida hiljem, nimelt aastal 1926, nimetas keemik Gilbert Lewis "footoniteks".

Igal footonil on energia, mida saab määrata valemiga E = hv – Plancki konstant korrutatuna kiirguse sagedusega.
Kooskõlas Max Plancki ideega sai seletatavaks nähtus, mille Hertz avastas 1887. aastal ja mida seejärel aastatel 1888–1890 põhjalikult uuris Stoletov. Aleksander Stoletov uuris eksperimentaalselt fotoelektrilist efekti ja kehtestas kolm fotoelektrilise efekti seadust (Stoletovi seadused):
Fotokatoodile langeva elektromagnetilise kiirguse konstantse spektraalse koostise korral on küllastusfotovool võrdeline katoodi energiavalgustusega (teisisõnu: katoodilt 1 sekundi jooksul välja löödud fotoelektronide arv on otseselt võrdeline kiirguse intensiivsusega) .
Fotoelektronide maksimaalne algkiirus ei sõltu langeva valguse intensiivsusest, vaid selle määrab ainult selle sagedus.
Igal ainel on fotoelektrilise efekti punane piir, st minimaalne valguse sagedus (olenevalt aine keemilisest olemusest ja pinna seisundist), millest allpool on fotoelektriline efekt võimatu.
Hiljem, 1905. aastal, täpsustas Einstein fotoelektrilise efekti teooriat. Ta näitab, kuidas valguse kvantteooria ning energia jäävuse ja muundamise seadus suurepäraselt seletavad seda, mis juhtub ja mida vaadeldakse. Einstein kirjutas üles fotoelektrilise efekti võrrandi, mille eest sai ta 1921. aastal Nobeli preemia:

Tööfunktsioon A on siin minimaalne töö, mida elektron peab tegema aine aatomist lahkumiseks. Teine liige on elektroni kineetiline energia pärast väljumist.
See tähendab, et footon neeldub aatomi elektronis, mille tõttu elektroni kineetiline energia aatomis suureneb neeldunud footoni energia võrra.
Osa sellest energiast kulub aatomist lahkuvale elektronile, elektron lahkub aatomist ja on võimeline vabalt liikuma. Ja suunas liikuvad elektronid pole midagi muud kui elektrivool või fotovool. Sellest tulenevalt saame rääkida EMF-i esinemisest aines fotoelektrilise efekti tulemusena.

See on, Päikesepatarei töötab tänu selles töötavale fotoelektrilisele efektile. Aga kuhu lähevad "välja löödud" elektronid fotogalvaanilises muunduris? Fotoelektriline muundur ehk päikesepatarei või fotoelement on seega fotoefekt selles ilmneb ebatavaliselt, see on sisemine fotoefekt ja sellel on isegi eriline nimetus “klapi fotoefekt”.
Päikesevalguse mõjul tekib pooljuhi p-n-siirdes fotoelektriline efekt ja tekib emf, kuid elektronid ei lahku fotoelemendist, kõik toimub blokeerivas kihis, kui elektronid lahkuvad ühest kehaosast, liikudes teise ossa. sellest.
Maakoores olev räni moodustab 30% selle massist, mistõttu seda kasutatakse kõikjal. Pooljuhtide eripära üldiselt seisneb selles, et nad ei ole juhid ega dielektrikud, nende juhtivus sõltub lisandite kontsentratsioonist, temperatuurist ja kokkupuutest kiirgusega.
Ribavahe pooljuhis on mitu elektronvolti ja see on just energiaerinevus aatomite valentsriba ülemise taseme, kust elektronid põgenevad, ja juhtivusriba alumise taseme vahel. Räni puhul on ribalaius 1,12 eV – just see, mida on vaja päikesekiirguse neelamiseks.

Niisiis, p-n ristmik. Fotoelemendis olevad legeeritud ränikihid moodustavad p-n ristmiku. Siin luuakse elektronidele energiabarjäär; nad lahkuvad valentsribast ja liiguvad ainult ühes suunas; augud liiguvad vastupidises suunas. Nii tekib päikesepatareis vool ehk päikesevalgusest toodetakse elektrit.
Footonitega kokkupuutunud Pn-liide ei lase laengukandjatel – elektronidel ja aukudel – liikuda muul viisil kui ühes suunas, need eralduvad ja satuvad barjääri vastaskülgedele. Ja olles ühendatud koormusahelaga ülemise ja alumise elektroodi kaudu, loob fotoelektriline muundur päikesevalguse käes välises vooluringis.
See elektritootmise meetod põhineb päikesevalgus, õpikutes nimetatud – Footonid. Meie jaoks on see huvitav, sest nii nagu liikuval õhuvoolul, on ka valgusvoolul energiat! Ühe astronoomilise ühiku (149 597 870,66 km) kaugusel Päikesest, kus asub meie Maa, on päikesekiirguse voo tihedus 1360 W/m2. Ja läbinud maa atmosfääri, kaotab vool peegelduse ja neeldumise tõttu oma intensiivsuse ning maapinnal on see juba ~ 1000 W/m2. Siit algab meie töö: kasutada valgusvoo energiat ja muuta see igapäevaelus vajalikuks energiaks – elektriliseks.

Selle teisenduse mõistatus leiab aset väikesel kaldnurkadega pseudoruudul, mis on lõigatud 125 mm läbimõõduga ränisilindrist (joonis 2) ja selle nimi on . Kuidas?
Sellele küsimusele said vastuse füüsikud, kes avastasid sellise nähtuse nagu fotoelektriline efekt. Fotoelektriline efekt on nähtus, kus elektronid paiskuvad valguse mõjul aine aatomitest välja.
Aastal 1900 Saksa füüsik Max Planck pakkus välja hüpoteesi: valgus kiirgab ja neeldub eraldi portsjonitena - kvantid(või footonid). Iga footoni energia määratakse järgmise valemiga: E =h∙ ν (tuhaakt) kus h- Plancki konstant on võrdne 6,626 × 10 -34 J∙s, ν - footoni sagedus. Plancki hüpotees selgitas fotoelektrilise efekti fenomeni, mille avastas 1887. aastal saksa teadlane Heinrich Hertz ja uuris eksperimentaalselt vene teadlane Aleksandr Grigorjevitš Stoletov, kes saavutas saadud tulemuste kokkuvõttel järgmise kolm fotoelektrilise efekti seadust:
- Valguse konstantse spektraalse koostise korral on küllastusvoolu tugevus otseselt võrdeline katoodile langeva valgusvooga.
- Valguse poolt väljutatavate elektronide esialgne kineetiline energia suureneb lineaarselt valguse sageduse suurenedes ega sõltu selle intensiivsusest.
- Fotoelektrilist efekti ei teki, kui valguse sagedus on väiksem kui igale ainele iseloomulik teatud väärtus, mida nimetatakse punaseks piiriks.
Fotoelektrilise efekti teooria, mis selgitab FEP-is valitsevat mõistatust, töötas välja Saksa teadlane Albert Einstein 1905. aastal, selgitades seadusi.  fotoelektriline efekt, kasutades valguse kvantteooriat. Tuginedes energia jäävuse ja muundamise seadusele, pani Einstein üles fotoelektrilise efekti käigus tekkiva energiabilansi võrrandi:
fotoelektriline efekt, kasutades valguse kvantteooriat. Tuginedes energia jäävuse ja muundamise seadusele, pani Einstein üles fotoelektrilise efekti käigus tekkiva energiabilansi võrrandi:
 Kus: h∙
ν
- footoni energia, A– tööfunktsioon – minimaalne töö, mis tuleb teha, et aine aatomist elektron lahkuks. Seega selgub, et valguse osake – footon – neeldub elektroni, mis omandab täiendava kineetilise energia ½m∙v 2
ja teostab aatomist lahkumise tööd, mis annab võimaluse vabalt liikuda. Ja elektrilaengute suunatud liikumine on elektrivool ehk õigemini öeldes aines tekib elektromotoorjõud – E.M.F.
Kus: h∙
ν
- footoni energia, A– tööfunktsioon – minimaalne töö, mis tuleb teha, et aine aatomist elektron lahkuks. Seega selgub, et valguse osake – footon – neeldub elektroni, mis omandab täiendava kineetilise energia ½m∙v 2
ja teostab aatomist lahkumise tööd, mis annab võimaluse vabalt liikuda. Ja elektrilaengute suunatud liikumine on elektrivool ehk õigemini öeldes aines tekib elektromotoorjõud – E.M.F.
Einstein pälvis 1921. aastal Nobeli preemia fotoelektrilise efekti võrrandi eest.
 Naastes minevikust tänapäeva, näeme, et päikesepatarei “süda” on FEP (pooljuhtfotoelement), milles realiseerub hämmastav looduse ime - Valve PhotoEffect (VPE). See seisneb elektromotoorjõu ilmnemises p-n-siirdes valguse mõjul. VFE või fotoelektriline efekt tõkkekihis, on nähtus, mille puhul elektronid lahkuvad kehast, minnes läbi liidese teise tahkesse kehasse (pooljuht).
Naastes minevikust tänapäeva, näeme, et päikesepatarei “süda” on FEP (pooljuhtfotoelement), milles realiseerub hämmastav looduse ime - Valve PhotoEffect (VPE). See seisneb elektromotoorjõu ilmnemises p-n-siirdes valguse mõjul. VFE või fotoelektriline efekt tõkkekihis, on nähtus, mille puhul elektronid lahkuvad kehast, minnes läbi liidese teise tahkesse kehasse (pooljuht).
Pooljuhid- need on materjalid, mis oma erijuhtivuse poolest asuvad vahepealsel positsioonil juhtide ja dielektrikute vahel ning erinevad juhtidest erijuhtivuse tugeva sõltuvuse poolest lisandite kontsentratsioonist, temperatuurist ja erinevat tüüpi kiirgusest. Pooljuhid on ained, mille ribavahemik on mitme elektronvoldi suurusjärgus [eV]. Ribavahe on elektronide energiate erinevus pooljuhtkristallides juhtivusriba alumise tasandi ja pooljuhi valentsriba ülemise tasandi vahel.
Pooljuhtide hulka kuulub palju keemilisi elemente: germaanium, räni, seleen, telluur, arseen jt, tohutul hulgal sulameid ja keemilisi ühendeid (galliumarseniid jne) Looduses levinuim pooljuht on räni,  moodustab umbes 30% maakoorest.
moodustab umbes 30% maakoorest.
Ränist pidi saama materjal selle laialdase looduses esinemise, kerguse ja sobiva 1,12 eV ribalaiuse tõttu päikesevalgusest energia neelamiseks. Tänapäeval on kaubanduslike maapealsete süsteemide turul kõige olulisemad kristalliline räni (umbes 90% maailmaturust) ja õhukese kilega päikesepatareid (umbes 10% turust).
 Kristallilise räni fotogalvaaniliste muundurite (PVC) disaini põhielement on p-n-siirde. Lihtsustatud kujul võib päikesepatarei kujutada "võileivana": see koosneb ränikihtidest, mis on legeeritud p-n-siirde moodustamiseks.
Kristallilise räni fotogalvaaniliste muundurite (PVC) disaini põhielement on p-n-siirde. Lihtsustatud kujul võib päikesepatarei kujutada "võileivana": see koosneb ränikihtidest, mis on legeeritud p-n-siirde moodustamiseks.
Pn-siirde üks peamisi omadusi on selle võime olla voolukandjatele energiabarjääriks, st võimaldada neil läbida ainult ühes suunas. Sellel mõjul põhineb elektrivoolu genereerimine päikesepatareides. Elemendi pinnale langev kiirgus tekitab pooljuhi mahus erineva märgiga laengukandjaid - elektrone (n) ja auke (p). Tänu oma omadustele pn-siirde "eraldab" need, võimaldades igal tüübil läbida ainult oma "oma" poole ning elemendi mahus kaootiliselt liikuvad laengukandjad satuvad tõkke vastaskülgedele, misjärel nad saab üle kanda välisesse vooluringi, et tekitada päikesepatareiga ühendatud suletud ahelas pinge ja elektrivool.
Fotosünteesi uurimise ajalugu ulatub 1771. aasta augustisse, mil inglise teoloog, filosoof ja amatöör-looduseteadlane Joseph Priestley (1733–1804) avastas, et taimed suudavad “korrigeerida” õhu omadusi, mis põlemise või selle koostist muutva või põlemise või selle koostise muutumise tagajärjel. loomade tegevus. Priestley näitas, et taimede juuresolekul muutub “riknenud” õhk taas sobivaks põlemiseks ja loomade elu toetamiseks.
Ingenhausi, Senebieri, Saussure'i, Boussingault' ja teiste teadlaste edasiste uuringute käigus leiti, et taimed valgustades eraldavad hapnikku ja neelavad õhust süsihappegaasi. Taimed sünteesivad orgaanilisi aineid süsihappegaasist ja veest. Seda protsessi nimetati fotosünteesiks.
Robert Mayer, kes avastas energia jäävuse seaduse, tegi 1845. aastal ettepaneku, et taimed muudavad päikesevalguse energia fotosünteesi käigus tekkinud keemiliste ühendite energiaks. Tema sõnul "püütakse kinni kosmoses levivad päikesekiired" ja säilitatakse vajaduse korral hilisemaks kasutamiseks. Seejärel uuris vene teadlane K.A. Timirjazev tõestas veenvalt, et kõige olulisemat rolli päikesevalguse energia kasutamisel taimede poolt mängivad rohelistes lehtedes esinevad klorofülli molekulid.
Fotosünteesi käigus tekkivaid süsivesikuid (suhkruid) kasutatakse energiaallikana ja ehitusmaterjalina erinevate orgaaniliste ühendite sünteesiks taimedes ja loomades. Kõrgemates taimedes toimuvad fotosünteesi protsessid kloroplastides, taimeraku spetsiaalsetes energiat muundavates organellides.
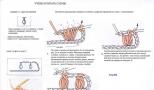
Kloroplasti skemaatiline kujutis on näidatud joonisel fig. 1.
Välis- ja sisemembraanidest koosneva kloroplasti topeltkesta all on laiendatud membraanstruktuurid, mis moodustavad suletud vesiikulid, mida nimetatakse tülakoidideks. Tülakoidmembraanid koosnevad kahest lipiidimolekulide kihist, mis sisaldavad makromolekulaarseid fotosünteetilisi valkude komplekse. Kõrgemate taimede kloroplastides on tülakoidid rühmitatud granaatideks, mis on lamestatud ja tihedalt kokku surutud kettakujuliste tülakoidide virnad. Grana üksikute tülakoidide jätkuks on nendest väljaulatuvad teradevahelised tülakoidid. Kloroplasti membraani ja tülakoidide vahelist ruumi nimetatakse stroomiks. Stroomas on kloroplasti molekulid RNA, DNA, ribosoomid, tärkliseterakesed, aga ka arvukalt ensüüme, sealhulgas neid, mis tagavad CO2 omastamise taimede poolt.
Väljaanne valmis Sushi E’xpress ettevõtte toel. Ettevõte "Sushi E'xpress" pakub Novosibirskis sushi kohaletoimetamise teenuseid. Tellides sushit firmast Sushi E’xpress, saad kiirelt kaasa maitsva ja tervisliku roa, mis on valmistatud professionaalsete kokkade poolt kõige värskematest ja kõrgeima kvaliteediga toorainetest. Sushi E’xpress firma kodulehte külastades saate tutvuda pakutavate rullide hindade ja koostisega, mis aitavad otsustada roa valiku üle. Sushi kohaletoimetamise tellimuse esitamiseks helista 239-55-87
Fotosünteesi heledad ja tumedad etapid
Kaasaegsete kontseptsioonide kohaselt on fotosüntees fotofüüsikaliste ja biokeemiliste protsesside jada, mille tulemusena taimed sünteesivad päikesevalguse energiat kasutades süsivesikuid (suhkruid). Fotosünteesi arvukad etapid jagunevad tavaliselt kaheks suureks protsessirühmaks – heledad ja tumedad faasid.
Fotosünteesi valgusetappe nimetatakse tavaliselt protsesside kogumiks, mille tulemusena sünteesitakse valgusenergia toimel adenosiintrifosfaadi (ATP) molekulid ja moodustub redutseeritud nikotii(NADP H), mis on kõrge kontsentratsiooniga ühend. väheneb potentsiaal, ilmneb. ATP molekulid toimivad rakus universaalse energiaallikana. Teadaolevalt kasutatakse ATP molekuli makroergiliste (st energiarikaste) fosfaatsidemete energiat enamikes energiat tarbivates biokeemilistes protsessides.
Fotosünteesi valgusprotsessid toimuvad tülakoidides, mille membraanid sisaldavad taimede fotosünteesiaparaadi põhikomponente - valgust koguvaid pigmendi-valgu ja elektronide transpordi komplekse, aga ka ATP süntaasi kompleksi, mis katalüüsib ATP moodustumist adenosiinist. difosfaat (ADP) ja anorgaaniline fosfaat (P i) (ADP + Ф i → ATP + H 2 O). Seega salvestub fotosünteesi valgusfaaside tulemusena taimedes neeldunud valguse energia ATP molekulide ja tugeva redutseerija NADP H kõrge energiaga keemiliste sidemete kujul, mida kasutatakse süsivesikute sünteesiks. fotosünteesi nn tumedad etapid.
Fotosünteesi pimedaid etappe nimetatakse tavaliselt biokeemiliste reaktsioonide kogumiks, mille tulemusena neeldub taimedesse atmosfääri süsihappegaas (CO 2) ja tekivad süsivesikud. Tumedate biokeemiliste transformatsioonide tsüklit, mis viib orgaaniliste ühendite sünteesini CO 2 -st ja veest, nimetatakse Calvin-Bensoni tsükliks, mis on nimetatud nende protsesside uurimisse otsustava panuse andnud autorite järgi. Erinevalt tülakoidmembraanis paiknevatest elektronide transpordi ja ATP süntaasi kompleksidest lahustuvad stroomas ensüümid, mis katalüüsivad fotosünteesi “tumedaid” reaktsioone. Kloroplasti membraani hävimisel uhutakse need ensüümid stroomast välja, mille tulemusena kaotavad kloroplastid süsihappegaasi neelamise võime.
Mitmete orgaaniliste ühendite muundumiste tulemusena Calvin-Bensoni tsüklis moodustub kloroplastides kolmest CO 2 molekulist ja veest glütseraldehüüd-3-fosfaadi molekul, mille keemiline valem on CHO-CHOH-CH 2 O–PO 3 2-. Sel juhul kulub ühe glütseraldehüüd-3-fosfaadis sisalduva CO 2 molekuli kohta kolm ATP molekuli ja kaks NADP H molekuli.
Orgaaniliste ühendite sünteesiks Calvin-Bensoni tsüklis ATP molekulide suure energiaga fosfaatsidemete hüdrolüüsireaktsiooni käigus eralduv energia (reaktsioon ATP + H 2 O → ADP + Ph i) ja NADP H tugev redutseerimispotentsiaal. kasutatakse molekule.Põhiosa kloroplastis tekkivatest molekulidest glütseraldehüüd-3-fosfaat siseneb taimeraku tsütosooli, kus see muundatakse fruktoos-6-fosfaadiks ja glükoos-6-fosfaadiks, millest edasiste transformatsioonide käigus moodustub suhkur. fosfaat, sahharoosi eelkäija. Tärklis sünteesitakse kloroplasti jäänud glütseraldehüüd-3-fosfaadi molekulidest.
Energia muundamine fotosünteesi reaktsioonikeskustes
Taimede, vetikate ja fotosünteetiliste bakterite fotosünteetilisi energiat konverteerivaid komplekse on hästi uuritud. Kindlaks on tehtud energiat muundavate valgukomplekside keemiline koostis ja ruumiline struktuur, selgitatud energia muundamise protsesside järjekord. Vaatamata fotosünteesiaparaadi koostise ja molekulaarstruktuuri erinevustele on kõigi fotosünteetiliste organismide fotoreaktsioonikeskustes energia muundamise protsesside üldised mustrid. Nii taimset kui ka bakteriaalset päritolu fotosünteesisüsteemides on fotosünteesiaparaadi ainus struktuurne ja funktsionaalne üksus fotosüsteem, mis sisaldab valgust koguvat antenni, fotokeemilist reaktsioonikeskust ja sellega seotud molekule – elektronide kandjaid.
Vaatleme esmalt kõikidele fotosünteesisüsteemidele iseloomulikke päikesevalguse energia muundamise üldpõhimõtteid ja seejärel peatume lähemalt fotoreaktsioonikeskuste ja kloroplastide elektronide transpordiahela toimimise näitel kõrgemates taimedes.
Valgust koguv antenn (valguse neeldumine, energia migratsioon reaktsioonikeskusesse)
Fotosünteesi kõige esimene elementaarne toiming on valguse neeldumine klorofülli molekulide või abipigmentide poolt, mis on osa spetsiaalsest pigmendi-valgu kompleksist, mida nimetatakse valguse kogumise antenniks. Valgust koguv antenn on makromolekulaarne kompleks, mis on loodud valguse tõhusaks püüdmiseks. Kloroplastides sisaldab antennikompleks suurel hulgal (kuni mitusada) klorofülli molekule ja teatud koguses valguga tihedalt seotud abipigmente (karotenoide).
Ereda päikesevalguse käes neelab üksik klorofülli molekul valguskvante suhteliselt harva, keskmiselt mitte rohkem kui 10 korda sekundis. Kuna aga klorofülli molekule on fotoreaktsioonikeskuse kohta suur hulk (200–400), aktiveerub reaktsioonikeskus ka suhteliselt nõrga valguse intensiivsusega lehele taimede varjutamise tingimustes üsna sageli. Valgust neelavate pigmentide ansambel toimib sisuliselt antennina, mis oma küllaltki suure suuruse tõttu tabab efektiivselt päikesevalgust ja suunab selle energia reaktsioonikeskusesse. Varju armastavatel taimedel on reeglina suurem valgust koguv antenn võrreldes kõrge valguse tingimustes kasvavate taimedega.
Taimedes on peamised valgust koguvad pigmendid klorofülli molekulid. a ja klorofüll b, neelab nähtavat valgust lainepikkusega λ ≤ 700–730 nm. Eraldatud klorofülli molekulid neelavad valgust ainult kahes suhteliselt kitsas päikesespektri ribas: lainepikkustel 660–680 nm (punane valgus) ja 430–450 nm (sinine-violetne valgus), mis muidugi piirab päikesevalguse kasutamise efektiivsust. kogu langeva päikesevalguse spekter rohelisele lehele.
Valgust koguva antenni poolt neeldunud valguse spektraalne koostis on aga tegelikult palju laiem. Seda seletatakse asjaoluga, et valgust koguva antenni osaks olevate klorofülli agregeeritud vormide neeldumisspekter nihkub pikemate lainepikkuste suunas. Koos klorofülliga sisaldab valgust koguv antenn abipigmente, mis suurendavad selle töö efektiivsust, kuna need neelavad valgust spektri nendes piirkondades, kus valgust koguva antenni peamine pigment klorofülli molekulid neelavad. valgus suhteliselt nõrgalt.
Taimedes on abipigmendid karotenoidid, mis neelavad valgust lainepikkuse piirkonnas λ ≈ 450–480 nm; fotosünteetiliste vetikate rakkudes on need punased ja sinised pigmendid: fükoerütriinid punavetikates (λ ≈ 495–565 nm) ja fükotsüaniinid sinivetikates (λ ≈ 550–615 nm).
Valguskvanti neeldumine klorofülli (Chl) molekuli või abipigmendi poolt viib selle ergutamiseni (elektron liigub kõrgemale energiatasemele):
Chl + hν → Chl*.
Ergastatud klorofülli molekuli Chl* energia kandub üle naaberpigmentide molekulidele, mis omakorda võivad selle üle kanda teistele valgust koguva antenni molekulidele:
Chl* + Chl → Chl + Chl*.
 Ergastusenergia võib seega migreeruda läbi pigmendimaatriksi, kuni ergastus jõuab lõpuks fotoreaktsioonikeskusesse P (selle protsessi skemaatiline esitus on näidatud joonisel 2):
Ergastusenergia võib seega migreeruda läbi pigmendimaatriksi, kuni ergastus jõuab lõpuks fotoreaktsioonikeskusesse P (selle protsessi skemaatiline esitus on näidatud joonisel 2):
Chl* + P → Chl + P*.
Pange tähele, et ergastatud olekus on klorofülli molekulide ja teiste pigmentide olemasolu väga lühike, τ ≈ 10 –10 –10 –9 s. Seetõttu on teatud tõenäosus, et teel reaktsioonikeskusesse P võib selliste lühiajaliste pigmentide ergastatud olekute energia kasutult kaduda – hajuda soojuseks või vabaneda valguskvanti kujul (fluorestsentsnähtus). Tegelikkuses on aga energia migratsiooni efektiivsus fotosünteesi reaktsioonikeskusesse väga kõrge. Kui reaktsioonikeskus on aktiivses olekus, on energiakao tõenäosus reeglina mitte suurem kui 10–15%. Päikeseenergia kasutamise kõrge efektiivsus tuleneb sellest, et valgust koguv antenn on väga korrastatud struktuur, mis tagab pigmentide väga hea interaktsiooni üksteisega. Tänu sellele saavutatakse valgust neelavate molekulide ergastusenergia kõrge ülekandekiirus fotoreaktsioonikeskusesse. Ergastusenergia ühelt pigmendilt teisele "hüppamise" keskmine aeg on reeglina τ ≈ 10 –12 –10 –11 s. Ergastuse kogumigratsiooniaeg reaktsioonikeskusesse ei ületa tavaliselt 10 –10 –10 –9 s.
Fotokeemiline reaktsioonikeskus (elektronide ülekanne, eraldunud laengute stabiliseerimine)
Kaasaegsetele ideedele reaktsioonikeskuse struktuuri ja fotosünteesi esmaste etappide mehhanismide kohta eelnesid A.A. Krasnovski, kes avastas, et elektronide doonorite ja aktseptorite juuresolekul on valguse poolt ergastatud klorofülli molekulid võimelised pöörduvalt redutseerima (aksepteerima elektroni) ja oksüdeeruma (loovutama elektroni). Seejärel avastasid Cock, Witt ja Duyzens taimedes, vetikates ja fotosünteetilistes bakterites spetsiaalsed klorofülli iseloomuga pigmendid, mida nimetatakse reaktsioonikeskusteks, mis oksüdeeruvad valguse toimel ja on tegelikult fotosünteesi ajal peamised elektronide doonorid.
Fotokeemiline reaktsioonikeskus P on spetsiaalne klorofülli molekulide paar (dimeer), mis toimib lõksuna valgust koguva antenni pigmendimaatriksis rändavale ergastusenergiale (joonis 2). Nii nagu vedelik voolab laia lehtri seintelt selle kitsasse kaela, suunatakse valguse energia, mida neelavad kõik valgust koguva antenni pigmendid, reaktsioonikeskusesse. Reaktsioonikeskuse ergastamine käivitab fotosünteesi käigus valgusenergia edasiste muundumiste ahela.
Pärast reaktsioonitsentri P ergastamist toimuvate protsesside jada ja fotosüsteemi energia vastavate muutuste diagramm on skemaatiliselt kujutatud joonisel fig. 3.
Koos klorofülli P dimeeriga sisaldab fotosünteesikompleks primaarsete ja sekundaarsete elektronide aktseptorite molekule, mida me tinglikult tähistame kui A ja B, ning ka primaarset elektronide doonorit, molekuli D. Ergastatud reaktsioonikeskusel P* on madal väärtus. afiinsus elektronide suhtes ja seetõttu loovutab see kergesti lähedalasuvale primaarsele elektronaktseptorile A:
D(P*A)B → D(P + A –)B.
Seega väga kiire (t ≈10–12 s) elektronide ülekande tulemusel P*-st A-sse realiseerub teine põhimõtteliselt oluline päikeseenergia muundamise etapp fotosünteesi käigus - laengu eraldumine reaktsioonikeskuses. Sel juhul moodustub tugev redutseerija A – (elektronidoonor) ja tugev oksüdeeriv aine P + (elektronide aktseptor).
Molekulid P + ja A – paiknevad membraanis asümmeetriliselt: kloroplastides paikneb reaktsioonikeskus P + tülakoidi sisekülje poole suunatud membraani pinnale lähemal ja aktseptor A – asub väljast lähemal. Seetõttu tekib fotoindutseeritud laengu eraldumise tulemusena membraanil elektripotentsiaalide erinevus. Valguse indutseeritud laengu eraldumine reaktsioonikeskuses on sarnane elektrilise potentsiaali erinevuse tekitamisega tavalises fotoelemendis. Siiski tuleb rõhutada, et erinevalt kõigist tehnoloogias tuntud ja laialdaselt kasutatavatest energia fotokonverteritest on fotosünteesi reaktsioonikeskuste tööefektiivsus väga kõrge. Laengu eraldamise efektiivsus aktiivsetes fotosünteesi reaktsioonikeskustes ületab reeglina 90–95% (päikesepatareide parimate näidete kasutegur ei ületa 30%).
Millised mehhanismid tagavad reaktsioonikeskustes nii kõrge energia muundamise efektiivsuse? Miks aktseptorile A üle kantud elektron ei pöördu tagasi positiivselt laetud oksüdeeritud tsentrisse P + ? Eraldatud laengute stabiliseerumine on tagatud peamiselt sekundaarsete elektronide transpordiprotsesside tõttu, mis järgnevad elektroni P*-st A-le ülekandmisel. Taastatud primaarsest aktseptorist A läheb elektron väga kiiresti (10–10–10–9 sekundiga) sekundaarne elektronaktseptor B:
D(P + A –)B → D(P + A)B – .
Sel juhul ei eemaldu mitte ainult elektron positiivselt laetud reaktsioonitsentrist P +, vaid ka kogu süsteemi energia väheneb märgatavalt (joonis 3). See tähendab, et elektroni vastassuunas ülekandmiseks (üleminek B – → A) peab ta ületama üsna kõrge energiabarjääri ΔE ≈ 0,3–0,4 eV, kus ΔE on energiatasemete erinevus kahe oleku vahel. süsteem, milles elektron on vastavalt kandjal A või B. Sel põhjusel kuluks elektroni tagasipöördumiseks redutseeritud molekulilt B - oksüdeeritud molekuli A - palju rohkem aega kui otseseks üleminekuks A - → B. Teisisõnu, edasisuunas kandub elektron palju kiiremini kui tagurpidi. Seetõttu väheneb pärast elektroni ülekandmist sekundaarsele aktseptorile B oluliselt selle tagasipöördumise ja positiivselt laetud “auguga” P + rekombineerumise tõenäosus.
Teine eraldatud laengute stabiliseerumist soodustav tegur on oksüdeeritud fotoreaktsioonikeskuse P + kiire neutraliseerimine tänu elektronidoonorilt D P +-le tarnitud elektronile:
D(P + A)B – → D + (PA)B – .
Olles saanud elektroni doonormolekulilt D ja naasnud algsesse redutseeritud olekusse P, ei suuda reaktsioonikeskus enam redutseeritud aktseptoritelt elektroni vastu võtta, kuid nüüd on ta valmis uuesti süttima - andma elektroni selle kõrval paiknev oksüdeerunud primaarne aktseptor A. See on sündmuste jada, mis toimub kõigi fotosünteesisüsteemide fotoreaktsioonikeskustes.
Kloroplasti elektronide transpordiahel
Kõrgemate taimede kloroplastides on kaks fotosüsteemi: fotosüsteem 1 (PS1) ja fotosüsteem 2 (PS2), mis erinevad valkude koostise, pigmentide ja optiliste omaduste poolest. Valgust koguv antenn FS1 neelab valgust lainepikkusega λ ≤ 700–730 nm ja FS2 valgust lainepikkusega λ ≤ 680–700 nm. PS1 ja PS2 reaktsioonitsentrite valgusega indutseeritud oksüdatsiooniga kaasneb nende pleegitamine, mida iseloomustavad muutused nende neeldumisspektris λ ≈ 700 ja 680 nm juures. Vastavalt nende optilistele omadustele nimetati PS1 ja PS2 reaktsioonitsentreid P 700 ja P 680.
Kaks fotosüsteemi on omavahel ühendatud elektronkandjate ahela kaudu (joonis 4). PS2 on PS1 jaoks elektronide allikas. Valgusalgatusega laengu eraldamine fotoreaktsioonikeskustes P 700 ja P 680 tagab elektroni ülekande PS2-s lagunenud veest lõplikule elektroni aktseptorile - NADP + molekulile. Kahte fotosüsteemi ühendav elektronide transpordiahel (ETC) sisaldab elektronide kandjatena plastokinooni molekule, eraldi elektronide transpordivalgu kompleksi (nn b/f kompleks) ja vees lahustuvat valku plastotsüaniini (P c). Joonisel fig. 4.
PS2-s kantakse ergastatud tsentrist P*680 elektron esmalt üle primaarsele aktseptor-feofetiinile (Phe) ja seejärel plastokinooni molekulile Q A, mis on tihedalt seotud ühe PS2 valguga,
Y(P* 680 Phe)Q A Q B → Y(P + 680 Phe –)Q A Q B → Y(P + 680 Phe)Q A – Q B .
Seejärel kantakse elektron üle teisele plastokinooni molekulile QB ja P 680 võtab vastu elektroni primaarselt elektrondoonorilt Y:
Y(P + 680 Phe)Q A – Q B → Y + (P 680 Phe)Q A Q B – .
 Plastokinooni molekul, mille keemiline valem ja asukoht lipiidide kahekihilises membraanis on näidatud joonisel fig. 5, on võimeline vastu võtma kahte elektroni. Pärast seda, kui PS2 reaktsioonikeskus süttib kaks korda, saab plastokinooni Q B molekul kaks elektroni:
Plastokinooni molekul, mille keemiline valem ja asukoht lipiidide kahekihilises membraanis on näidatud joonisel fig. 5, on võimeline vastu võtma kahte elektroni. Pärast seda, kui PS2 reaktsioonikeskus süttib kaks korda, saab plastokinooni Q B molekul kaks elektroni:
Q B + 2е – → Q B 2– .
Negatiivselt laetud Q B 2- molekulil on kõrge afiinsus vesinikioonide suhtes, mida ta püüab kinni stroomaruumist. Pärast redutseeritud plastokinooni Q B 2– (Q B 2– + 2H + → QH 2) protoneerimist moodustub selle molekuli elektriliselt neutraalne vorm QH 2, mida nimetatakse plastokinooliks (joonis 5). Plastokinool toimib kahe elektroni ja kahe prootoni liikuva kandjana: pärast PS2-st lahkumist saab QH 2 molekul tülakoidmembraani sees hõlpsasti liikuda, tagades PS2 ühenduse teiste elektronide transpordikompleksidega.
Oksüdeeritud reaktsioonitsenter PS2 R 680 on erakordselt kõrge elektronafiinsusega, s.t. on väga tugev oksüdeerija. Tänu sellele lagundab PS2 vett, mis on keemiliselt stabiilne ühend. PS2 osaks olev veejaotuskompleks (WSC) sisaldab oma aktiivses keskuses rühma mangaaniioone (Mn 2+), mis toimivad P680 elektronide doonoritena. Annetades elektrone oksüdeeritud reaktsioonikeskusele, muutuvad mangaaniioonid positiivsete laengute "akumulaatoriteks", mis on otseselt seotud vee oksüdatsioonireaktsiooniga. P 680 reaktsioonitsentri järjestikuse neljakordse aktiveerimise tulemusena koguneb VRC Mn sisaldavasse aktiivtsentri neli tugevat oksüdatiivset ekvivalenti (ehk neli auku) oksüdeeritud mangaaniioonide (Mn 4+) kujul, mis , interakteerudes kahe veemolekuliga, katalüüsivad lagunemisreaktsiooni vett:
2Mn 4+ + 2H2O → 2Mn 2+ + 4H + + O2.
Seega toimub pärast nelja elektroni järjestikust ülekandmist VRC-st P 680-le korraga kahe veemolekuli sünkroonne lagunemine, millega kaasneb ühe hapnikumolekuli ja nelja vesinikiooni vabanemine, mis sisenevad kloroplasti intratülakoidi ruumi.
PS2 toimimise käigus tekkinud QH 2 plastokinooli molekul difundeerub tülakoidmembraani lipiidide kaksikkihti b/f kompleksi (joonis 4 ja 5). Kui see puutub kokku b/f kompleksiga, seondub QH 2 molekul sellega ja kannab seejärel kaks elektroni üle. Sel juhul vabaneb iga b/f kompleksi poolt oksüdeeritud plastokinooli molekuli kohta tülakoidi sees kaks vesinikiooni. Kompleks b/f toimib omakorda elektronidoonorina plastotsüaniinile (P c), mis on suhteliselt väike vees lahustuv valk, mille aktiivne keskus sisaldab vase iooni (plastotsüaniini redutseerimis- ja oksüdatsioonireaktsioonidega kaasnevad muutused plastotsüaniini valentsis). vase ioon Cu 2+ + e – ↔ Cu+). Plastotsüaniin toimib lülina b/f kompleksi ja PS1 vahel. Plastotsüaniini molekul liigub kiiresti tülakoidi sees, tagades elektronide ülekande b/f kompleksist PS1-le. Redutseeritud plastotsüaniinist läheb elektron otse PS1 – P 700 + oksüdeerunud reaktsioonikeskustesse (vt joonis 4). Seega kandub PS1 ja PS2 koostoime tulemusena PS2-s lagunenud veemolekulist kaks elektroni lõpuks elektronide transpordiahela kaudu NADP + molekuli, tagades tugeva redutseerija NADP H moodustumise.
Miks vajavad kloroplastid kahte fotosüsteemi? Teatavasti toimivad ühe fotosüsteemiga edukalt fotosünteesivad bakterid, mis kasutavad elektronidoonorina oksüdeerunud reaktsioonikeskuste taastamiseks erinevaid orgaanilisi ja anorgaanilisi ühendeid (näiteks H 2 S). Kahe fotosüsteemi ilmumine on suure tõenäosusega tingitud asjaolust, et ühe nähtava valguse kvanti energiast ei piisa, et tagada vee lagunemine ja elektroni efektiivne läbimine mööda kandjamolekulide ahelat veest NADP +-ni. Umbes 3 miljardit aastat tagasi ilmusid Maale sinivetikad ehk tsüanobakterid, mis omandasid võime kasutada vett elektronide allikana süsinikdioksiidi vähendamiseks. Praegu arvatakse, et PS1 pärineb rohelistest bakteritest ja PS2 lilladest bakteritest. Pärast seda, kui evolutsiooniprotsessi käigus oli PS2 koos PS1-ga "kaasatud" ühte elektronide ülekandeahelasse, sai võimalikuks energiaprobleemi lahendamine - ületada hapniku/vee paaride ja NADP + / üsna suur erinevus redokspotentsiaalide vahel. NADP H. Vett oksüdeerima võimeliste fotosünteetiliste organismide esilekerkimine sai üheks olulisemaks etapiks eluslooduse arengus Maal. Esiteks on vetikad ja rohelised taimed, olles "õppinud" vett oksüdeerima, omandanud ammendamatu elektronide allika NADP + redutseerimiseks. Teiseks täitsid nad vett lagundades Maa atmosfääri molekulaarse hapnikuga, luues nii tingimused organismide kiireks evolutsiooniliseks arenguks, mille energia on seotud aeroobse hingamisega.
Elektronide transpordiprotsesside sidumine prootoniülekandega ja ATP süntees kloroplastides
Tavaliselt kaasneb elektronide ülekandega ETC kaudu energia vähenemine. Seda protsessi võib võrrelda keha spontaanse liikumisega mööda kaldtasandit. Elektroni energiataseme langus tema liikumisel mööda ETC-d ei tähenda sugugi seda, et elektronide ülekanne on alati energeetiliselt kasutu protsess. Tavalistes kloroplasti funktsioneerimise tingimustes ei lähe suurem osa elektronide transpordil vabanevast energiast asjatult raisku, vaid seda kasutatakse spetsiaalse energiat konverteeriva kompleksi, mida nimetatakse ATP süntaasiks, tööks. See kompleks katalüüsib energeetiliselt ebasoodsat ATP moodustumise protsessi ADP-st ja anorgaanilisest fosfaadist P i (reaktsioon ADP + P i → ATP + H 2 O). Sellega seoses on tavaks öelda, et elektronide transpordi energiat loovutavad protsessid on seotud ATP sünteesi energiaaktseptori protsessidega.
Olulisemat rolli energia sidumise tagamisel tülakoidmembraanides, nagu ka kõigis teistes energiat muundavates organellides (mitokondrid, fotosünteetiliste bakterite kromatofoorid), mängivad prootoni transpordiprotsessid. ATP süntees on tihedalt seotud kolme prootoni ülekandega tülakoididest (3H +) strooma (3H välja +) ATP süntaasi kaudu:
ADP + Ф i + 3H sisse + → ATP + H 2 O + 3H välja + .
See protsess saab võimalikuks, kuna kandjate asümmeetrilise paigutuse tõttu membraanis viib kloroplastide ETC toimimine tülakoidi sees liigse prootonite kogunemiseni: vesinikioonid neelduvad väljastpoolt NADP etappides. + redutseerimine ja plastokinoolide moodustumine ning vabanevad tülakoidide sees vee lagunemise ja plastokinooli oksüdatsiooni faasis (joon. 4). Kloroplastide valgustamine toob kaasa märkimisväärse (100–1000-kordse) vesinikioonide kontsentratsiooni suurenemise tülakoidide sees.
Niisiis oleme vaadelnud sündmuste ahelat, mille käigus päikesevalguse energiat salvestatakse kõrge energiasisaldusega keemiliste ühendite - ATP ja NADP H - energia kujul. Neid fotosünteesi valgusfaasi tooteid kasutatakse pimedas faasis. moodustada süsihappegaasist ja veest orgaanilisi ühendeid (süsivesikuid). ATP ja NADP H moodustumiseni viivad energia muundamise peamised etapid hõlmavad järgmisi protsesse: 1) valgusenergia neeldumine valgust koguva antenni pigmentide poolt; 2) ergastusenergia ülekandmine fotoreaktsioonikeskusesse; 3) fotoreaktsioonikeskuse oksüdeerimine ja eraldunud laengute stabiliseerimine; 4) elektronide ülekanne mööda elektronide transpordiahelat, NADP H teke; 5) vesinikioonide transmembraanne ülekanne; 6) ATP süntees.
1. Alberts B., Bray D., Lewis J., Roberts K., Watson J. Rakkude molekulaarbioloogia. T. 1. – M.: Mir, 1994. 2. trükk.
2. Kukushkin A.K., Tihhonov A.N. Loengud taimede fotosünteesi biofüüsikast. – M.: Moskva Riikliku Ülikooli kirjastus, 1988.
3. Nichols D.D. Bioenergia. Sissejuhatus kemosmootilise teooriasse. – M.: Mir, 1985.
4. Skulatšev V.P. Bioloogiliste membraanide energia. – M.: Nauka, 1989.